
Electroacupuncture-Induced Central Nervous System Activation Promotes Mesenchymal Stem Cell Release

Introduction
Acupuncture is an ancient therapy that works by inserting needles into specific body points known as acupoints. Electroacupuncture (EA) enhances this technique by adding electrical stimulation. Acupoints are found in areas with lower electrical resistance and higher conductivity, due to neural and vascular features in the skin. Histological studies show that these points are rich in nerve endings, small blood vessels, lymphatics, and mast cells. Some acupoints align with particular neural structures like nerves and nerve networks.
Acupuncture’s pain-relieving effects are mainly thought to come from releasing opioid peptides in the central nervous system (CNS) when sensory pathways are activated. Adenosine also plays a role, and blocking its metabolism can extend the benefits of acupuncture.
Despite knowing about the anatomical and signaling properties of acupoints, the full mechanisms behind acupuncture's systemic benefits and healing effects are not completely understood. This study aims to explore how EA affects both the central and peripheral nervous systems using two sets of acupoints (1: LI-4, LI-11, GV-14, Bai-hui; 2: ST-36, LIV-3, GV-14, and Bai-hui; Supporting Information Fig. S1), which are linked to treating arthritis, asthma, and immune modulation.
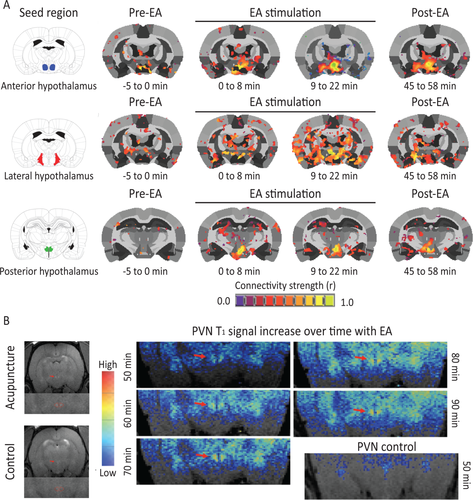 Figure 1
Figure 1
Electroacupuncture (EA) stimulation increases hypothalamic functional connectivity in rats. (A): Rat brains were monitored through functional magnetic resonance imaging during administration of EA. Functional connectivity increased within the hypothalamus and between the hypothalamus and adjacent brain region with progression of treatment (n = 6). (B): A representative example of the increase in signal (over time) in the PVN of a rat receiving acupuncture (n = 1). Abbreviations: EA, electroacupuncture; PVN, paraventricular nucleus.
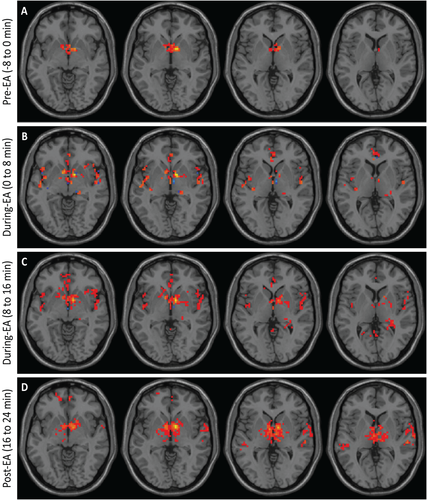 Figure 2
Figure 2
Electroacupuncture (EA) stimulation increases hypothalamic functional connectivity in humans. Brains from normal subjects were monitored using functional magnetic resonance imaging before, during, and after EA. Functional connectivity increased within the hypothalamus and between the hypothalamus and adjacent brain regions with progression of treatment (n = 6).
As these regions have been associated with hematopoietic stem cell mobilization 6, peripheral blood was examined in the rat before and following EA for evidence of mesenchymal stem cell (MSC) mobilization. A 313% increase in the circulating MSC population, Lin-CD90+CD44HI cells, was detected in the blood of EA-treated rats 2 hours post-EA compared with baseline (Fig. 3A, 3B).
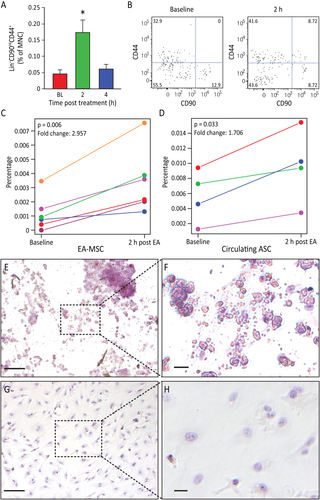 Figure 3
Figure 3
Electroacupuncture (EA) stimulation induced mesenchymal stem cell (MSC) mobilization. (A): Rat peripheral blood MSC were increased (p = .0063) after EA. Circulating MSC were defined as Lin-(CD45-CD31-erythroid-CD11b-) cells that were positive for CD44 and CD90. Gated cells increased post treatment (n = 11 for baseline and 4 hours, n = 9 for 2 hours). (B): Representative flow charts for rat Lin- cells are shown at baseline and 2 hours samples. (C): The percentage of human peripheral blood MSC increased in post EA-treatment (p = .006, n = 6). (D): The percentage of circulating MSCs from adipose tissue (AD-MSC) is significantly elevated 2 hours post EA-treatment (p = .033, n = 4). (E, F): EA-mobilized MSCs were expanded in vitro. After undergoing adipogenesis differentiation, EA-mobilized MSCs developed fat deposits as seen by Oil Red staining, which were not seen in the undifferentiated control cells (G, H). Magnification bars = (E, G): 100 µm; (F, H): 50 µm. Abbreviations: ASC, adipose stem cells; EA, electroacupuncture; MNC, mononuclear cell.
Characterization of Human EA Mobilized MSC as Adipocyte-Derived MSC
The availability of human antibodies to fully characterize the populations of cells mobilized by EA led us to examine these populations in the human subjects who underwent fMRI. Due to limitations with the fMRI equipment, only points GV-14, LI-11, and LI-14 could be used in this experiment. We observed an increase in MSC in the peripheral blood 2 hours after EA in all subjects (Fig. 3C), while total lymphocyte numbers remained unchanged. To further characterize the MSC population and determine if they were released from adipose stores, we examined the levels of CD34+CD45-CD31- cells. Two hours after EA, CD34+CD45-CD31- cells were increased in four out of six individuals (Fig. 3D), and the response was proportional to the body mass index (BMI) of the subjects, as previously noted. Two individuals with a BMI of less than 18.5 did not show an increase in this population. Human EA-mobilized MSC were expanded in vitro and underwent adipogenic differentiation. Adipogenesis potential was confirmed by the cells’ ability to form lipid droplets, as established through oil red O staining (Fig. 3E–3H).
Since hypothalamic activation (Fig. 1B) preceded the mobilization of circulating MSC, we proposed that EA-induced connectivity changes contributed to the subsequent release of these cells into the peripheral blood. Exogenous administration of epinephrine or dopamine in rats resulted in a similar increase in the Lin-CD90HICD44+ cell population in the circulation (Supporting Information Fig. S2A–S2C), supporting the role of the CNS in mobilizing MSC and in EA activating these key CNS centers.
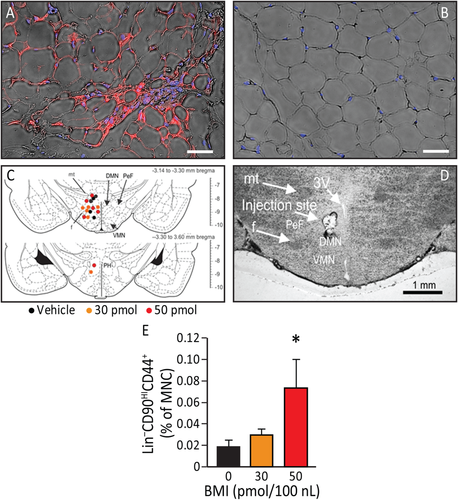 Figure 4
Figure 4
Electroacupuncture (EA) increases sympathetic activation leading to browning of white adipose tissue and the effects of EA can be duplicated by pharmacological disinhibition of hypothalamus (sympathetic activation). (A, B): UCP1 immunofluorescence (red) detectable in inguinal subcutaneous adipose tissue (blue: adipocytes nuclei) from animals that underwent EA treatment (A) but not in control (B). EA resulted in an increase in beige adipocytes (n = 4). Magnification bars = 50 µm. (C): The effects of EA can be duplicated by pharmacological disinhibition of hypothalamus. Rats underwent injection of either vehicle, 30 pmol or 50 pmol/100 nL of the GABAA receptor antagonist bicuculline methiodide. Sites of injections are represented on a coronal section from the tuberal hypothalamus from a Standard Steroetaxic Atlas of the Rat brain 11. Colored circles indicate injection sites (black, orange, and red represent vehicle, 30 pmol and 50 pmol, respectively). (D): Representative photomicrograph showing an injection site from one rat. Magnification bar = 1 mm. (E): There was a significant increase (p = .027) in Lin-CD90HICD44+ cells in the plasma 4 hours post injection (n = 6). Data presented as means ± SEM. Abbreviations: BMI, bicuculline methiodide; DMN, dorsomedial hypothalamic nucleus; f, fornix; MNC, mononuclear cells; mt, mammillothalamic tract; PeF, perifornical hypothalamus; PH, posterior hypothalamic nucleus; VMN, ventromedial hypothalamic nucleus; 3V, third ventricle.
Pharmacological Disinhibition of the Dorsomedial Regions of the Tuberal Hypothalamus Mobilizes Lin-CD90HICD44+ Cells into the Circulation
To confirm that activation of the sympathetic nervous system (SNS) can mediate the release of MSCs, we performed stereotaxic injections of the GABAA receptor antagonist bicuculline methiodide (BMI) into the dorsomedial regions of the tuberal hypothalamus to disinhibit these regions. Histological verification of the injection sites is shown in the illustration (Fig. 4C). The exact locations of all injection sites are indicated on coronal sections from a Standard Stereotaxic Atlas of the Rat Brain (Fig. 4C) and a photomicrograph showing the representative injection site (Fig. 4D). In rats, BMI at a dose of 50 pmol increased the percentage of circulating Lin-CD90HICD44+ cells (F(2, 14) = 6.7, p = 0.027) at 4 hours post-injection of BMI (Fig. 4E) in the absence of EA. These results demonstrate that circulating Lin-CD90HICD44+ cells are mobilized during hypothalamic disinhibition and EA. However, the delay in release (4 hours) compared to EA (2 hours) may suggest an alternative mechanism of release.
EA-Treated Rodents Exhibit Reduced Mechanical Hyperalgesia, Increased Serum Interleukin-10 Levels, and Enhanced Tissue Remodeling Following Partial Achilles Tendon Rupture
To address the potential anti-inflammatory and analgesic effects of EA-mobilized circulating MSCs, we analyzed the impact of an EA treatment regimen on injury-induced hyperalgesia in rats, both with and without the β-blocker propranolol. Treatments included EA (i.e., applied to forelimb points LI-4, LI-11, GV-14, and Bai-hui, and hindlimb points ST36, LIV-3, GV-14, and Bai-hui in horses), sham EA (i.e., applied to skin not associated with an acupoint), or EA plus propranolol (i.e., a potential inhibitor of the sympathetic effects of EA). EA or sham EA was administered every other day for 2 weeks following injury. Using sham EA applied to non-immune acupoints or EA plus propranolol, nociceptive behavior elicited by von Frey mechanical stimulation did not change over time in the hind paw ipsilateral to the injury (Supporting Information Fig. S3). In contrast, mechanical hyperalgesia (assessed at both day 7 and 14) was significantly reduced (i.e., increased tolerance to pressure) in injured rodents subjected to EA at LI-4, LI-11, GV-14, and Bai-hui (Fig. 5A). At early stages of tendon repair, granulation tissues mainly synthesize type III collagen, while at later stages, intrinsic fibroblasts produce type I collagen, whose fibers are oriented longitudinally to replace type III collagen. At 14 days post-injury, EA sham-treated or EA plus propranolol did not alter type I collagen content, while type I collagen was significantly increased by EA (Fig. 5B); there was no change in type III collagen across treatments (Fig. 5C). These data suggest that EA may enhance the replacement of thinner and immature type III collagen fibers with mature type I collagen fibers in the injured tendon, thereby supporting better quality regeneration and tissue reorganization. Given that EA can reduce mechanical hyperalgesia and enhance tissue reorganization, we examined possible changes in the anti-inflammatory cytokine interleukin (IL)-10 in blood plasma. Previous evidence suggested that EA following surgical trauma contributes to increased levels of IL-10 by T-cells, and focal microinjections of IL-10 can diminish mechanical hyperalgesia. The results here demonstrate that tendon injury in rodents subjected to sham EA or EA combined with propranolol did not produce detectable changes in plasma IL-10. However, injured rodents treated with EA showed a significant increase in plasma IL-10. These results indicate that EA increases the production of the endogenous anti-inflammatory cytokine IL-10 (Fig. 5D).
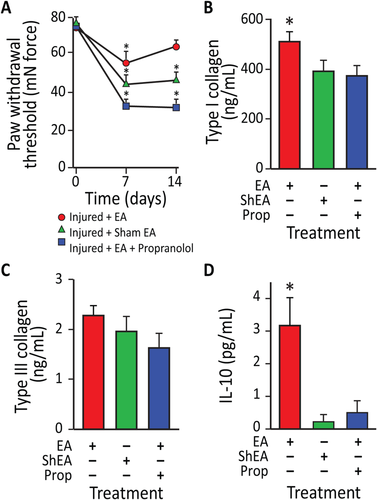 Figure 5
Figure 5
Electroacupuncture (EA)-treated rodents exhibit reduced mechanical hyperalgesia, enhanced tissue remodeling, and increased serum IL-10 levels following partial Achilles tendon rupture. (A): Effects of EA application on mechanical allodynia in rats at 7 and 14 days after partial tendon rupture in the right hind leg. Mechanical hypersensitivity was determined by measuring the change in weight-bearing forces on the affected limb. Behavioral changes in the hind paw tactile threshold (in millinewtons, mN) were observed in the hindpaw ipsilateral to the tendon injury 18 hours after EA treatment, EA sham treatment or EA treatment with propranolol. (*, p < .01 versus baseline. EA: n = 9; EA: sham n = 7; EA + propranolol: n = 10). (B): EA increased type I collagen content in injured tendons in rats at 14 days after unilateral Achilles tendon partial tenotomy. In EA-treated animals (n = 6), type-I collagen content was 24% greater in injured tendons than EA sham treated tendons (n = 7; p < .05) and 28% greater in injured tendons than EA + propranolol treated tendons (n = 8; p < .02). (C): In contrast, there was no difference in type-III collagen content between injured EA and EA sham or EA + propranolol treated tendons (p = .67). (D): IL-10 serum levels were also elevated in EA-treated rats compared with EA sham and EA + propanolol animals (p = .0041). Data presented as means ± SEM. Abbreviations: EA, electroacupuncture; IL-10, interleukin-10.
EA Performed over Immune Points in Pirt-GCaMP3 Mice Rapidly Activates Primary Sensory Neurons
We next examined mice to confirm that EA stimulation of acupoints (LI-4, LI-11, GV-14, and Bai-hui) resulted in the mobilization of MSCs. At 4 hours post-EA, murine MSCs, defined as Lin-PDGFRa+Sca-1+ cells, were significantly increased (p = 0.01) in peripheral blood. This response was markedly reduced when mice were pretreated with propranolol (Supporting Information Fig. S4). Immune acupoints are present on both the front and hind limbs of mice and are used based on their accessibility in the species undergoing EA. Thus, we used acupoints in both front limbs (LI-4, LI-11) and hind limbs (ST-36 and LIV-3). To distinguish the afferent stimulation of the hypothalamus by peripheral acupoints, we treated Pirt-GCaMP3 mice with either a noxious hind paw pinch (100g force) or EA directed at the hind limb acupoints (Fig. 6A–6F) [15]. Hind paw stimulation evoked robust and transient increases in 10–20 neurons per DRG in naïve mice, averaging 12.7 ± 3.0 per DRG (Fig. 6A, 6B), nearly all of which were small diameter neurons (<20 µm; Supporting Information Fig. S5; Supporting Information Video 1), suggestive of pain fibers. Placement of acupuncture needles alone did not elicit activity in sensory neurons (Fig. 6C, 6D; Supporting Information Video 2). We observed a striking pattern of neuronal activation in the DRG following EA stimulation; many activated neurons were medium to large diameter neurons (medium [20–25 µm]; large [>25 µm]) with an average of 6.1 ± 4.0 neurons per ganglion (Fig. 6E, 6F; Supporting Information Video 3), suggesting activation of touch fibers.
EA at Immune Points in the Forelimbs Preferentially Mobilizes MSCs, Whereas EA of Immune Points in the Hind Limb Mobilizes Macrophages into the Circulation
Acupuncture in horses and humans is pursued with the same rigor and is not as technically difficult as acupuncture in rodents. We examined the forelimb (LI-4, LI-11, GV-14, and Bai-hui) and hind limb immune points (ST36, LIV-3, GV-14, and Bai-hui) in horses. Peripheral blood from horses (n = 30) undergoing EA at LI-4, LI-11, GV-14, and Bai-hui was examined first. Due to the limited availability of equine MSC antibodies compared to humans and mice, stem cell mobilization into peripheral blood was confirmed by measuring circulating cell colony-forming ability in vitro. While colony-forming cells were rarely seen at baseline, colony-forming ability was easily detected in blood samples obtained 2 and 4 hours after EA (Fig. 7A), the same time points examined in humans and rats. Blood collected at 2 and 4 hours post-EA using mock acupoints approximately 1 cm from the immune points in the same horses did not give rise to colonies in vitro. Importantly, and representing a more critical control than simply sham acupoints, the use of metabolic points similarly did not yield significantly more colonies in vitro (Fig. 7A). To further verify the stem/progenitor characteristics of the equine cells, clonogenic potential was determined using single-cell assays. EA-mobilized cells showed robust clonogenic potential, with over 75% proliferating into two or more cells, and over 50% forming large colonies of 10,000 cells or more; these levels of proliferation are generally reflective of stem/progenitor cells (Fig. 7B). The MSC origin of the mobilized colony-forming cells was confirmed by their in vitro differentiation into osteocytes, as demonstrated by positive staining for calcium deposits (Fig. 7C, panels a, b [controls: panels e, f]), as well as their adipogenic differentiation (Fig. 7C, panels c, d [controls: panels g, h]) and chondrogenic differentiation (Fig. 7C, panels i, j), supporting that equine EA-mobilized cells display MSC lineage characteristics. When the EA-mobilized equine cells were examined in the in vivo angiogenesis assay, the cells did not directly form blood vessels (Fig. 7D, panel a) or lumenize, thus supporting a nonendothelial origin. However, co-implantation of the EA-mobilized equine cells with human cord blood-derived endothelial colony-forming cells (ECFCs) significantly enhanced ECFC vasculogenesis (Fig. 7D, panels b, c) and increased the number of human vessels with an arterial morphology (Fig. 7E). Furthermore, when the EA-mobilized MSCs were co-cultured with human ECFCs (hECFCs) in vitro, a significant increase in HEY2 expression was observed in the endothelial cells, indicating that the addition of the equine cells promoted arteriogenesis, as the Notch signaling pathway is known to be active in arterial vascular endothelial cells (Fig. 7F). Overall, these data support that EA-mobilized cells display MSC characteristics and enhance hECFC vasculogenesis and arteriogenesis.
To compare EA-mobilized MSCs (EA-MSCs) with other MSC populations derived from either adipose stem cells (ASC) or bone marrow (BM-MSC), we performed gene array studies. Of the ~30,000 genes present on the EquGene-1_0-st GeneChip, 678 showed significant differences between EA-MSC and BM-MSC, 1,164 between EA-MSC and ASC, and 1,193 between ASC and BM-MSC (all p < 0.05 and absolute fold change >2) (Supporting Information Tables S1–S3). Principal component analysis mapping (Supporting Information Fig. S6A), hierarchical clustering (Supporting Information Fig. S6B), and partitioning clustering (Supporting Information Fig. S6C) showed that EA-MSCs, BM-MSCs, and ASCs segregated into distinct groups. This suggests that the EA-mobilized MSC population may be derived from a source distinct from either adipose tissue or bone marrow, or that their mobilization into the systemic circulation modified their gene expression compared to BM-MSCs or ASCs obtained directly from their tissue sources. Genes specifically upregulated in EA-MSCs compared with BM-MSCs and ASCs (Supporting Information Fig. S6C, Cluster 1) encoded numerous proteins involved in cell cycle control and progression, DNA replication and repair, endothelial cell physiology, and adhesion and migration (BGN, CTH, DHFR, ENG, EDN1, MYOF, PROCR, VEGF, several integrins, and SERPINB2). Additionally, this group included genes coding for enzymes implicated in extracellular matrix synthesis, such as proteoglycans (e.g., HAS2, possibly involved in vasculogenesis, CHSY1, GCNT4) and collagens (COL1A1, COL1A2, COL3A1, COL5A1, COL5A2, and COL12A1). Furthermore, EA-MSCs expressed several growth hormones, hormone receptors, and members of their signaling pathways (FGF5, BDNF, HTR2A, ADORA2B, and RLN) (Supporting Information Tables S1 and S3).
Genes showing the greatest decreases in EA-MSCs (Supporting Information Fig. S6C, Cluster 2) were acute-phase response genes and protease inhibitors (HP, SAA1, JAM2, C1S, C1R, and SLPI), consistent with the fact that EA point stimulation reduces acute and chronic inflammation (Supporting Information Tables S2, S4).
The pathways involved with the genes differentially expressed between EA-MSCs and BM-MSCs were analyzed using Ingenuity Pathway Analysis software (IPA) and included cellular growth and proliferation, hepatic pathways and embryonic stem cell pluripotency, DNA damage response, axonal guidance signaling, cardiovascular system development, mitotic roles of polo-like kinase, cell cycle: G2/M DNA damage checkpoint regulation, cell cycle control of chromosomal replication, GADD45 signaling, and ATM signaling (Supporting Information Table S5).
In contrast, BM-MSCs demonstrated increased expression of genes involved in inflammatory responses (acute phase, cytokine signaling), cell motility, and response to hormones and growth factors (Supporting Information Fig. S6C, Cluster 3), while ASCs displayed highly increased expression of genes related to cholesterol, fatty acids, and lipid metabolism, inflammatory response, and redox homeostasis (Supporting Information Fig. S6C, Cluster 4).
Changes in representative genes (ADAM23, COL1A1, ENG, FGF5, GCNT4, HP, IGFBP3, RLN, SAA1, and SERPINB2) were further validated by real-time polymerase chain reaction (Supporting Information Fig. S7).
Contemporary interpretation of EA must consider contributions from both dry needling and electrical stimulation, as performed in clinical applications such as transcutaneous electrical stimulation (TENS). Therefore, we next investigated whether TENS performed with adhesive electrodes versus EA at immune acupoints LI-4, LI-11, GV-14, and Bai-hui would result in similar
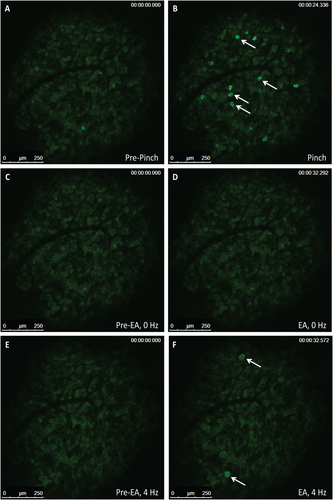 Figure 6
Figure 6
Small diameter primary afferent sensory neurons show sensitivity to pinch stimulus and medium-large diameter neurons show sensitivity to electroacupuncture (EA) in Pirt-GCaMP3 mice with intact dorsal root ganglia (DRG). Representative fluorescent GCaMP3 neuronal imaging response by lumbar DRG in vivo before (A) and after a mechanical press of the hindpaw using a 100g force (B). The mechanical force evokes a robust fluorescent GCaMP3 Ca2+ response in numerous small sensory neurons (B; white arrows). Acupuncture needles alone failed to elicit neuronal changes in the same lumbar DRG of the Pirt-GCamp3 mouse (C, D). EA-stimulation of the ST-36 and LIV-3 accupoints produced rapid activation of medium–larger diameter of the lumbar DRG (E, F; white arrows) (n = 4, p < .0001). Abbreviation: EA, electroacupuncture.
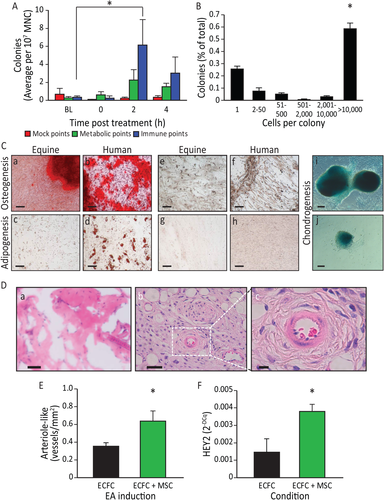 Figure 7
Figure 7
Electroacupuncture (EA)-mediated sympathetic stimulation induces mesenchymal stem cell (MSC) release into the circulation. (A): EA mobilized cells are highly proliferative and potentiate vasculogenesis. Equine peripheral blood mononuclear cells (MNC) showed an increased colony-forming ability 2 hours (p < .05) post administration of EA at immune points (n = 7), while cells obtained from the same horses when they underwent mock treatment or treatment at metabolic points did not. (B): The EA-mobilized cells demonstrated high proliferative capacity, when plated in a single-cell assay, with over 50% proliferating into large colonies (p < .001 vs. all groups). (C): Equine peripheral blood MNCs were cultured to the third passage and then differentiated into key mesenchymal lineages. (a): Following culture with osteogenic induction media, the mobilized equine cells showed strong osteogenic potency, demonstrated by Alizarin red staining of calcium deposits (red: Alizarin red). (b): Human mesenchymal cells responded in a similar fashion when cultured under identical conditions. Equine cells when cultured under control media (1:1 Ham's F12 and low glucose Dulbecco's modified Eagle's medium, 15% fetal bovine serum) (e) and human cells under control conditions (f) did not show Alizarin red staining. (c): The EA-mobilized equine cells showed a weak adipogenic response, demonstrated by oil red O staining of lipid deposits (red: oil red O) when cultured under adipogenic conditions; (d) human MSC showed a much stronger response than the equine cells when cultured under identical adipogenic conditions. Under control conditions, neither equine MSCs (g) or human MSCs (h) showed oil red O staining. (i, j) When cells were cultured under chondrogenesis differentiation media they were able to differentiate into chondrogenic lineages, demonstrated by Alcian Blue staining of proteoglycans in the cell masses. Magnification bars = 50 µm. (D): In vivo angiogenesis assay. (a): When equine cells were incorporated into a three-dimensional type I pig skin collagen plug and placed under the skin of NOD/SCID mice no capillaries were formed. (b): Human endothelial colony forming cells (hECFC) were inserted into the porcine collagen plugs together with equine MSC, and implanted into the flank of NOD/SCID mice. (c): A higher magnification of the boxed area in (b), showing a bona fide blood vessel. Magnification bars = (a, b): 50 µm; (c): 10 µm. (E): When quantified, the hECFC-MSC had a significant increase of arteriogenesis compared with hECFC alone (p = .02, n = 5 for ECFCs alone, n = 7 for combined hECFC-MSC group). (F): After 48 hours in vitro, cells were isolated, total mRNA was extracted and HEY2 expression levels were quantified by quantitative real-time polymerase chain reaction. HEY2 was elevated in the mixed cell treatment when compared with ECFC alone (p = .006, n = 4). All data presented as means ± SEM. Abbreviations: EA, electroacupuncture; ECFC, endothelial colony forming cell; MNC, mononuclear cell; MSC, mesenchymal stem cell.
Discussion
In this study, we make the highly novel observation that EA at immune points releases key populations into the peripheral blood (MSC and macrophages) that can modulate physiological responses to injury. While the effect of EA on analgesia is well-accepted, the effect of EA on the release of reparative cell populations is largely unknown. Equally important is that the effect of EA we observed is consistent across four species. fMRI studies in both humans and rats support that EA activates the hypothalamus, leading to the mobilization of MSC. These cells can be ex vivo expanded and demonstrate the phenotypic and functional characteristics of MSC. Depending on the set of immune points used, the reparative population released varies. When EA is performed at GV-14 and Bai-hui with forelimb points (LI-4 and LI-11), MSC are released into the blood. Conversely, EA at GV-14 and Bai-hui with hind limb points (ST-36 and LIV-3) results in the release of macrophage-like cells (Supporting Information Fig. S1). Similar responses to those observed with forelimb immune points were also seen with injections of either epinephrine or dopamine and with pharmacological disinhibition of the tuberal hypothalamus using the GABAA receptor antagonist bicuculline methiodide in rats. Pharmacological disinhibition of the tuberal hypothalamus increases plasma concentrations of norepinephrine by 100% and epinephrine by 400% [16]. Deep brain stimulation of the tuberal hypothalamus in humans also enhances sympatho-excitation [17]. Furthermore, these electrical activities could be replicated by TENS stimulation at the precise EA points. Thus, the mobilization of MSCs is likely centrally mediated through hypothalamic activation and subsequent SNS activity. There is growing interest and evidence supporting the role of the SNS as a regulator of immune cell release, as appropriate activation and signaling are necessary to mobilize these cells into the bloodstream [6].
TENS is a widely accepted method of pain relief [18]. The use of TENS over acupoints is a relatively new application, sometimes referred to as transcutaneous EA [19]. Methods to mobilize circulating MSC (e.g., epinephrine, dopamine, substance P, and GM-CSF) or hematopoietic stem cells (e.g., chemotherapy, growth factors, or AMD3100 [20-23]) into the bloodstream are often associated with adverse side effects. However, the use of EA or TENS represents a simple and cost-effective approach to increasing MSC numbers in the peripheral blood, facilitating MSC harvest and ex vivo expansion.
EA has been reported to have long-lasting and powerful analgesic effects in models of both acute and chronic pain [24, 25]. Adrenergic receptors, GABAergic modulation through peripheral, spinal, and supraspinal mechanisms, and adenosine A1 receptors have been implicated in the underlying mechanisms of this effect [4, 26-28]. EA-induced mobilization of circulating MSC may directly or indirectly modulate anti-inflammatory and immunomodulatory properties in vivo [29, 30], as evidenced by increased serum IL-10 levels and reduced mechanical hyperalgesia. This suggests that EA limits the production of nociceptive proinflammatory cytokines and enhances tissue remodeling following tendon injury [31-33]. In contrast, chronic treatment with the non-selective β-adrenergic receptor antagonist propranolol effectively blocks EA's anti-inflammatory activity [34, 35]. Monocyte mobilization into peripheral blood can also be mediated by sympathetic signaling [36]. In our studies, the use of hind-limb points mobilized a cell type that acquired a macrophage-like phenotype in culture and demonstrated strong phagocytic activity in vitro. These activated cells could be playing a role in modulating the cytokine milieu, leading to EA-mediated repair and analgesia (Supporting Information Fig. S10). The specificity of cell mobilization according to the points warrants further study.
The activation of the SNS with EA is further supported by our results demonstrating that EA at the specific points LI-11, LI-4, GV-14, and Bai-hui promoted the browning of WAT in rats. Brown adipose tissue counteracts WAT function [37, 38]. This effect was achieved using an every-other-day EA protocol for 14 days. Shen et al. [39] used points typically associated with the treatment of obesity (acupoints ST-36 and ST-44) six times per week for 5 weeks in DIO mice to achieve a similar effect. Although they did not observe weight loss or a decrease in appetite in EA-treated mice, they did see a reduction in the ratio of WAT weight to body weight, suggesting that EA increased lipolysis in obese mice. WAT “browning” is driven by SNS stimuli, such as cold temperature, and signal transduction cascades triggered by catecholamines activating β3 adrenergic receptors [37, 41, 42]. UCP1+ thermogenic brown-like (beige/bright) adipocytes within WAT [43, 44] activate energy expenditure and can counteract the metabolic consequences of obesity [45, 46]. Even a mild reduction in lipid content in WAT associated with its browning activates energy expenditure and has positive metabolic benefits [47, 48]. Fat browning has been considered a promising avenue in diabetes treatment [49].
Conclusion
Acupuncture is among the oldest healing practices in the world and is currently one of the most rapidly growing complementary therapies. Our studies provide strong support for the use of EA at specific immune points to stimulate MSC and macrophage release into peripheral blood through hypothalamic and SNS activation. EA may facilitate tissue repair following injury by increasing the levels of circulating MSC and could be used to treat acute or chronic conditions associated with inflammation [49, 50]. Furthermore, fMRI and direct neurostimulation studies have confirmed SNS activation and the metabolically beneficial response of browning of WAT. The importance of adiposity is demonstrated in individuals with a body mass index greater than 18.5. We observed an increase in the specific subset of MSC, the ASC population, only in these individuals. Importantly, EA-stimulated “browning” of WAT can enhance metabolism and influence glucose sensitivity. Additionally, harvesting MSC from the blood of EA-treated human subjects and ex vivo expansion is feasible and may serve as a practical method for autologous cell therapy, avoiding the risks and discomfort associated with current, more invasive and toxic collection methods.
Acknowledgments
We thank the Angio BioCore at Indiana University School of Medicine for their work in the human studies, the Flow Cytometry Resource Facility at Indiana University Simon Cancer Center (partially funded by National Cancer Institute Grant P30. CA082709), the OSUCCC Microarray Shared Resource, where the equine GeneChips were processed, and Christopher Brown for his original illustration of the graphical abstract. This work was supported by NIH Grants R01EY012601-15, R01HL11070-03, R01DK090730-04, and R01EY007739-23 (to M.B.G.), U54 DK106846-01 and R01 HL109602 (to M.C.Y.), PR151924 (to M.E.B.), and DK100905, R01DK100905, and 101BX002209 (to F.A.W.), by the St. Vincent Foundation (to F.A.W.), and by the Cryptic Masons’ Medical Research Foundation (to K.L.M. and D.T.).
Author Contributions
T.E.S. and M.R.R.: study design, acquisition of data, data analysis and interpretation, manuscript writing, final approval of manuscript; E.B. and J.C.: study design, acquisition of data, data analysis and interpretation, manuscript writing; M.S.R., Y.D., A.B., V.J., J.A.S., A.L.B., D.O.T., S.D., B.M.D., S.L.C., S.D.F., R.K.F., S.J.W., and T.M.K.: acquisition of data; J.G.: study design, acquisition of data; Y.K., Y.Y., L.M.C.-P., E.S., J.A.M., and P.L.J.: acquisition of data, data analysis and interpretation; L.M. and K.A.: acquisition of data, data analysis and interpretation, manuscript writing; S.M.G.: data analysis and interpretation; K.L.M.: study design; M.S.K.: study design, acquisition of data, data analysis and interpretation; M.E.B.: manuscript writing; J.T.: data acquisition, data analysis and interpretation, manuscript writing; A.S.: data analysis and interpretation; M.F., M.G.K., and F.A.W.: study design, data acquisition, data analysis and interpretation, manuscript writing; L.J.C., S.L., and X.D.: study design, data acquisition, data analysis and interpretation; Z.G.: data acquisition; J.M.: data acquisition and analysis; H.X.: study design, data acquisition, manuscript writing; M.C.Y.: study design, data analysis and interpretation, manuscript writing, final approval of manuscript; M.B.G.: conception of ideas, study design, data acquisition, data analysis and interpretation, manuscript writing, final approval of manuscript. T.E.S. and M.R.R. are first authors who contributed equally to this article. F.A.W., H.X., M.C.Y., and M.B.G. are senior authors who contributed equally to this article.
Aug 26,2024